El núcleo es el organelo más
grande de la célula, en el se encuentra casi todo el acido desoxirribonucleico
(ADN), los componentes para la síntesis de ácido ribonucleico (ARN), y en su
nucleolo se ensamblan las subunidades ribosómicas. A grandes rasgos el núcleo
alberga
·
La cromatina: Ósea el material genético de la
célula
·
El nucleolo: Donde se forma el ARN ribosómico
·
El nucleoplasma: que contiene macromoléculas y
partículas nucleares que participan en el mantenimiento de la célula
 |
| Estructura General del nucleo |
En la mayoría de las veces el
núcleo suele tener una forma esférica, y estar ubicado en el centro de la
célula, pero en ocasiones su ubicación y forma suele ser distinta por lo que se
han ideado las siguientes clasificaciones para el mismo:
·
Por su posición: En esta clasificación se
divide en Polarizada o No Polarizada dependiendo si la célula se encuentra
sobre una lamina basal o no. (cuando hablemos de epitelio trataremos más a
detalle sobre la lámina basal)
Ø Polarizada:
Los núcleos que entran en está clasificación son aquellos cuyas células se
encuentran fijas a una lámina basal, por lo que los núcleos pueden ser:
Centrales si se ubican en el centro de la célula, Parabasales si se ubican
fuera del centro, o Basales si se encuentran cercanos (casi pegados) a la
lámina basal.
Ø No
Polarizada: Los núcleos que entran en esta clasificación son aquellos cuyas
células no están fijas a una lámina basal, por lo que los núcleos pueden ser: Centrales
si se ubican en el centro de la misma, Excéntrico si se encuentra fuera del
centro o Periférico si se encuentra en la periferia de la célula cercano (casi
pegado) a la membrana plasmática
·
Por la Forma del Núcleo: Este puede ser:
Esférico como en la mayoría de las células, Ahusado (Aplastado), o Lobulado
(Deforme)
 |
| Nucleos por su forma: 1 Defforme, 2 y4 Esferico, 3 Aplastado |
·
Por el numero de núcleos: En este caso las
células pueden ser Anucleada (Sin núcleo), Uninucleada (1 núcleo), Binucleada
(2 núcleos), Multinucleada (Más de 2 núcleos)
 |
| Nucleos por numero: 1 Anucleadas, 2 Uninucleadas, 3 Binucleadas, 4 Multinucleadas |
Envoltura Nuclear
El núcleo esta rodeado por la
envoltura nuclear que está formada por dos membranas, la membrana nuclear
interna y la membrana nuclear externa, ambas son paralelas
concéntricas separadas entre si por un espacio denominado cisterna perinuclear.
La envoltura nuclear está perforada por los poros nucleares que permiten
la comunicación entre el citoplasma y el núcleo. En estos poros ambas membranas
son continuas entre sí.
Membrana Nuclear Interna
Se encuentra en contacto
directo con el contenido nuclear y con las laminas nucleares, que son
una malla entrelazada de filamentos intermedios formada por laminas A, B1,
B2, y C, las cuales están situadas en la periferia del
nucleoplasma. Estas laminas ayudan a mantener el revestimiento de la membrana
nuclear interna gracias a su unión a proteínas integrales de la membrana como
los polipéptidos asociados a la lamina y la emerina. Ayudan en la
organización y soporte de la bicapa lipídica de la membrana y la cromatina
perinuclear, participan en la formación de complejos de poro nuclear y en el
ensamblaje de vesículas para volver a formar la envoltura nuclear al finalizar
la división celular (Proceso que dejaremos para otro momento).
Membrana Nuclear Externa
Contacta con el citoplasma y
continua con el Retículo Endoplásmico Rugoso (del que se hablara en la
siguiente entrada de este blog), Su superficie esta cubierta por una malla de
filamentos intermedios, la vimetina, y ribosomas que producen proteínas
destinadas a las membranas interna y externa.
Poros Nucleares
Los poros son interrupciones
de la envoltura nuclear donde la membrana interna y externa se fusionan dando
lugar a sitios donde el núcleo y el citoplasma se comunican. Gracias al
microscopio electrónico se ha visto que el poro está rodeado por estructuras no
membranosas que están incrustadas en su borde, las cuales forman el complejo
de poro nuclear que regula de forma selectiva el paso a través del poro.
 |
| Poros Nucleares vistos con Microscopio Electrónico |
Complejos de Poro Nuclear
Están formados por proteínas
conocidas como nucleoporinas acomodadas en tres estructuras en forma de anillo
octagonal (anillo citoplasmático, anillo radial luminal y anillo nuclear)
apilados uno encima de otro unidos entre sí por proteínas denominadas radiales,
también el complejo tiene fibras citoplasmáticos, tapón central y cresta
nuclear.
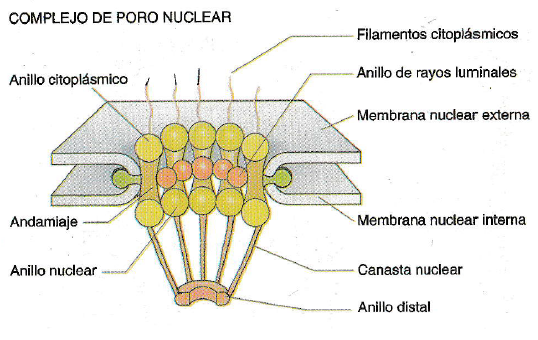

El Anillo Citoplasmático está
compuesto por ocho subunidades que bordean la cara citoplasmática del poro,
cada subunidad tiene un filamento citoplasmático, una proteína que se
une a Ran (una proteína que se une al guanosin trifosfato (GTP)), La función de
los filamentos citoplasmáticos es la de regular la entrada al núcleo moviendo
sustratos a lo largo de su longitud hacia el centro del poro.
El Anillo Radial Luminal está
formado por ocho subunidades que se proyectan hacia la luz del poro y la
cisterna perinuclear, estas proteínas se unen a las glicoproteínas del complejo
del poro nuclear. En el centro del anillo se encuentra el tapón central o
transportador que se acopla a las proteínas radiales, no se conoce su
función ya que no se sabe si es un tapón verdadero que limita la
difusión a través del poro o si es solo una carga atrapada durante el tránsito
en el poro.
El anillo nuclear es análogo
al anillo citoplasmático, colabora con la exportación de varios tipos de ARN.
En el anillo se encuentra la cresta nuclear que está suspendida del anillo
y sobresale hacia el nucleoplasma y se deforma durante la exportación nuclear,
unido a la cresta nuclear se encuentra el anillo distal.
Los poros nucleares tienen un
tamaño lo suficientemente grande como para permitir el paso de cualquier
sustancia por difusión simple, sin embargo, ese espacio en su mayoría es
ocupado por los componentes del complejo de poro nuclear, quedando espacio
únicamente para el paso para iones o moléculas pequeñas, por lo que las
moléculas grandes se transportan por un proceso de transporte mediado por
receptor. Algunos lugares de unión del complejo de poro nuclear deben reconocer
secuencias señal de las moléculas que den ser transportadas.
El trafico bidireccional entre
el núcleo y el citoplasma es mediado por un grupo de proteínas que contienen
señales de localización nuclear (NLS) conocidas como importinas, y señales de
exportación nuclear conocidas como exportinas (o PTAC por transportinas y
proteínas fijadoras de Ran).
Las exportinas transportan
moléculas del núcleo al citoplasma (ejemplo ARN), y las importinas hacen lo
contrario al introducir moléculas del citoplasma al núcleo (ejemplo subunidades
ribosómicas). El transporte de exportina y la Importina depende de un grupo de
proteínas fijadoras de GTP llamadas Ran que junto a otras nucleoporinas
facilitan la importación y exportación mediada por señales.
 |
| Funciones de RAN |
Cromatina
El ADN representa el material genético
de la célula se encuentra en el núcleo en forma de cromosomas, los cuales se
encuentran desenrollados en forma de cromatina. La cromatina es el complejo formado por ADN y
proteínas que representa a los cromosomas sin enrollar, De acuerdo con la
actividad transcripcional de la célula se puede encontrar condensada como
heterocromatina o extendida como eucromatina. La Heterocromatina es una
forma inactiva y condensada de la cromatina, mientras que la Eucromatina
representa una forma activa cuando las moléculas del ADN se Transcriben a ARN.
Ácidos Nucleicos
Los ácidos nucleicos ADN y ARN
son polinucleótidos que codifican la información genética usada para construir y
mantener a los organismos vivos. El ADN bicatenario es usado para dirigir los procesos
celulares. Las células convierten las instrucciones operativas del ADN en la secuencia
de nucleótidos de las moléculas de ARN monocatenario. Las moléculas de ARN
tienen múltiples funciones que incluyen la: síntesis de proteínas, regulación
de la expresión genética (control de si se sintetiza un polipéptido específico
y cuándo sucede) y protección contra los ácidos nucleicos introducidos en las
infecciones virales.
ADN
EI ADN es un polímero de nucleótidos
(un nucleótido es un compuesto químico por una base nitrogenada (Timina,
Adenina, Guanina o Citosina), un azúcar y una molécula de ácido fosfórico) unidos
covalentemente por medio de enlaces fosfodiéster 3'~
5'. Los enlaces fosfodiéster
unen el grupo 3'-hidroxilo de la desoxipentosa de un nucleótido al grupo
5'-hidroxilo de la desoxipentosa de un nucleótido adyacente a través de un
grupo fosfato. La larga cadena, no ramificada, resultante tiene polaridad, con
un extremo 5' (el extremo con el fosfato libre) y un extremo 3' (el extrema con
el hidroxilo libre) que no están unidos a otros nucleótidos. El ADN puede ser
monocatenario (es decir de una sola hebra como en los virus) o bicatenario formando
dobles hélices (como en la mayoría de eucariotas) o estructuras lineales y
circulares (Como en las bacterias).
 |
| Enlaces fosfodiester |
La Doble Hélice y el
Emparejamiento de Bases
En la doble hélice, las dos
cadenas se enrollan alrededor de un eje común Llamado eje de simetría. Las cadenas
están emparejadas de una manera antiparalela, es decir, el extremo 5' de una
hebra esta emparejado con el extremo 3' de la otra hebra. En la hélice de ADN,
el esqueleto hidrófilo de desoxirribosa-fosfato de cada cadena está en la parte
externa de la molécula mientras que las bases hidrófobas están apiladas en la
parte interna. Este acomodo es muy parecido a una escalera de caracol y crea
dos surcos uno mayor y otro menor Estos surcos proporcionan acceso y
posibilitan la unión de las proteínas reguladoras a sus secuencias de
reconocimiento específicas a lo largo de la cadena del ADN.
 |
| Doble Helice |
Correlación Clínica Ciertos antineoplásicos, como la dactinomicina
(actinomicina D), ejercen su efecto citotóxico intercalándose en el surco
estrecho de la doble hélice del ADN e interfiriendo así en la síntesis del ADN
y del ARN
Las bases de una Hebra de ADN
se emparejan con las de la otra a través de puentes de hidrogeno quedando
siempre unida adenina con una timidina por 2 puentes de hidrogeno y una citosina
con una guanina por 3 puentes de hidrogeno. De esta forma una hebra es
complementaria con la otra siguiendo la ley de Chargaff que dice que la
cantidad de adenina en las 2 hebras es proporcional a la de timidina.
 |
| Puentes de Hidrogeno y Emparejamiento de Bases |
Replicación del ADN
Aunque este tema es abordado
de forma más detallada por la biología molecular, aquí daremos un breve resumen
para que el lector tenga una pequeña noción del proceso y comprenda mejor los
siguientes temas, así mismo cabe aclarar que a pesar de que el proceso se rige
por los mismos principios en eucariotas y procariotas, existen diferencias
notables en cuanto a enzimas y ciertos pasos en el proceso. Dado que este blog
está enfocado a tejidos humanos únicamente nos centraremos en el proceso de los
eucariotas. Si el lector desea conocer el proceso en procariotas puede
consultar cualquier libro o sitio dedicado a la biología molecular o la bioquímica.
El proceso comienza con el
ensamblaje del complejo de replicación de preiniciación (preRC). El
ensamblaje del preRC comienza cuando el complejo de origen de replicación (ORC)
recluta a MCM una enzima con función de helicasa, es decir que rompe los
puentes de hidrogeno separando ambas hebras de ADN las cuales luego son
estabilizadas por proteínas de replicación A (RPA) que evitan que las hebras
se vuelvan a unir, de esta manera se forma la horquilla de replicación. Posteriormente
la enzima ADN Pol α (O ADN Primasa para los cuates) inicia la formación
de un ARN cebador y un pequeño fragmento de ADN de 10 a 20 nucleótidos
de longitud que es complementario al de la cadena principal, después las
enzimas ADN Pol δ y ADN Pol ε Continúan copiando la hebra. La replicación
de una hebra se da en sentido 3'~ 5', y la otra en 5'~ 3'. Pero sin embargo las
polimerasas solo pueden trabajar en sentido 3'~ 5', por lo que una de
las hebras se forma de manera continua, por lo que es denominada cadena
adelantada, en la que una sola ADN Pol ε copia toda la cadena de la horquilla,
y otra de manera discontinua denominada cadena retrasada ya que varias
ADN Pol δ copian la cadena en sentido 3'~ 5' dejando espacios denominados
fragmentos de Okazaki. Finalmente, las Enzimas FEN1 Dna2 retiran los
cebadores para que luego estos huecos sean llenados por enzimas ADN Ligasas
dando como resultado 2 ADN bicatenarios de doble hebra.
Alginas cosas que se deben
tomar en cuenta es que la replicación es semiconservativa, eso quiere decir que
cada nuevo ADN resultado de la replicación tiene una hebra de la molécula
original de ADN, y que el proceso se realiza en varias horquillas de
replicación a lo largo de la doble hélice que después se juntan.
Organización del ADN Eucariota
La longitud del ADN de una
sola célula humana seria mas de 1 metro por lo que este tiene que compactarse
demasiado para poder entrar en un núcleo celular para ello el ADN se une a una
serie de proteínas llamadas histonas que son proteínas que a un pH fisiológico
tienen carga positiva debido a su contenido de lisina y arginina que se unen
bien al ADN con carga negativa. Existen varios tipos de histonas que son H1, H2A,
H2B, H3 y H4. Las histonas y el ADN se unen entre si para formar una estructura
llamada nucleosoma, compuesta por un núcleo de 2 histonas de cada tipo excepto
de H1, y ADN, a su vez los nucleosomas están unidos entre si por ADN conector
unido a la histona H1. Varios nucleosomas forman polinucleosomas que adopta una
forma en espiral denominada fibra. La fibra se encuentra en bucles anclados por
varias proteínas que en conjunto darán lugar a los cromosomas el nivel de
compactación máximo del ADN.
 |
| Nucleosoma |
 |
| Organización del genoma eucariota |
El numero de cromosomas de
cada célula depende de la especie, en el ser humano por ejemplo son 46 o 23
pares de los cuales un miembro de cada par deriva de la madre y el otro del
padre. 22 de los 23 pare son denominados autosomas y el par restante es
denominado sexual ya que define el sexo del individuo desde la concepción. Las mujeres
poseen 2 cromosomas X mientras que el hombre 1 X y 1 Y.
ARN
Al igual que el ADN el ARN es
un polímero de nucleótidos unidos por enlaces fosfodiéster, pero con
diferencias en su estructura y su función. En primer lugar, el ARN es mas
pequeño que el ADN, sus nucleótidos tienen ribosa en lugar de desoxirribosa, el
ARN en comparación con el ADN solo es monocatenario, capaz de plegarse y formar
estructuras, y su diferencia más notoria es su carencia de timina, que es
remplazada en su lugar por uracilo como base nitrogenada. Existen 3 tipos
distintos de ARN que son:
·
ARN Ribosómico (ARNr): El cual se asocia con
varias proteínas para formar los ribosomas (De los que hablaremos en la
siguiente entrada de este blog). Para las células eucariotas existen cuatro
tipos 28S, 5.8S y 5S que forman la subunidad grande y 18S que forma la
subunidad pequeña. (S por coeficiente de sedimentación de Svedberg)
·
ARN de Transferencia (ARNt): son los
responsables de aportar los aminoácidos durante la síntesis de proteínas, por
lo que existe uno especifico para cada uno de los aminoácidos.
·
ARN Mensajero (ARNm): Transporta la información
genética del ADN nuclear al ribosoma donde sirve como un molde para la síntesis
de proteínas. Además de las regiones codificantes posee en el extremo 5´ una
caperuza de metil guanosina y en el 3´ una cola de Poliadenilato o Poli A que
evita la degradación del mismo por nucleasas al salir al citoplasma.
Síntesis de ARN o
Transcripción
El proceso en el cual se
produce una hebra de ARN a partir de una de las hebras del ADN de doble hélice
se le conoce como transcripción. El producto es enviado al ribosoma para ser
codificado en una proteína. Al igual que en la Replicación del ADN este tema es
tratado a detalle por la biología molecular por lo que aquí solo se hará una
breve explicación, al ser diferente el proceso en eucariotas y procariotas y al
ser este un blog enfocado a tejidos humanos nos centraremos en el proceso de
los eucariotas. Si el lector desea conocer más detalles o el proceso en
procariotas puede hacerlo en cualquier libro de biología molecular o bioquímica.
El ADN que será transcrito se
encuentra en la eucromatina y los que no serán transcritos son parte de la
heterocromatina, el cambio de la heterocromatina a la eucromatina es la remodelación
de la cromatina. Para ello el ADN debe separarse de las histonas a través de la
acetilación de sus lisinas por la enzima histona acetiltransferasa (HAT)
y la vuelta a la normalidad se da al retirar el acetilo por la histona
desacetilasa (HDAC).
La primer fase es denominada INICIACIÖN.
La enzima ARN Pol II es la enzima clave para la síntesis del ARNm a
partir de una hebra de ADN, pero para ello requiere la intervención de factores
de transcripción para acercar a la ADN Pol II a una secuencia promotora
denominada Caja TATA. Dichos factores de transcripción son TFIID que
reconoce a la caja TATA y se une a ella, TFIIF que acerca a la ADN Pol
II a la caja TATA y el TFIIH que cumple una función de helicasa para desenrollar
el ADN. Además de la ARN Pol II, otras polimerasas son la ARN Pol I que
sintetiza el ARNr, y la ARN Pol III que sintetiza el ARNt.
La siguiente fase la denomina ELONGACIÖN.
En está fase la ARN Pol II lleva a
cabo su trabajo copiando una hebra de ADN en ARN, En el caso de los ARNm las
ribonucleasas lo parten en varios ARN de varios tamaños y luego modificados por
exonucleasas, para los ARNt se eliminan secuencias de ambos extremos, y se
añade una CCA por la nucleotidiltransferasa, y para el ARNm se añade la cola de
poli A y la Caperuza CAP.
En el ultimo paso MADURACIÖN,
El ARN es eliminado de intrones, es decir regiones transcritas de ADN que
no codificaran proteínas, conservando únicamente los exones, que son los
transcritos que si codificaran proteínas y estos son empalmados entre si por
corte y empalme.
Nucleoplasma
El nucleoplasma es una
sustancia que rodea a los cromosomas y el nucleolo compuesta por gránulos de
intercromatina, pericromatina, ribonucleoproteínas (RNP) y la matriz nuclear.
Los granos de intercromatina
tienen RNP y varias enzimas como ATPasas, GTPasas, NAD pirofosfatasas entre
otras, aunque su función aun no es clara. Los granulos de pericromatina tienen
particulas de ribonucleoproteínas nucleares pequeñas (snRNP), de las cuales aun
no se sabe su función. La matriz por otra parte posee la mayor parte del ADN y
ARN y RNP residuales.
Nucléolo
Es una estructura nuclear no
delimitada por membrana alguna, se encarga de la síntesis del ARNr y el
ensamblaje de las subunidades ribosómicas. Se le han descrito 4 regiones que
son.
·
Centro Fibrilar: Que contienen ADN inactivo
·
Parte Fibrilar: Donde se transcribe ADN en ARNr
·
Parte Granulosa: Donde se ensamblan los
ribosomas
·
Matriz Nucleolar: Una red de fibras que
mantienen al nucléolo
En conclusión, el núcleo es
una estructura importante para la célula, ya que además de almacenar la
información genética lleva acabo los primeros pasos en el procesamiento y
trasmisión de la misma como lo es la replicación y transcripción, pero está
historia aún no acaba ya que falta la ultima parte del proceso llamada
traducción, pero por hoy ya fue demasiado así que lo dejaremos para otro día. Así
que no te pierdas nuestra próxima entrada donde hablaremos de los Ribosomas y
El Retículo Endoplásmico Rugoso y por supuesto la traducción.
Fuentes
Gartner, L., 2017. Texto De Histología Atlas A Color. 4th ed. Barcelona: Elsevier
Harvey, R., Ferrier, D. Palacios Martínez, R., 2011. Bioquímica. 5th ed. Philadelphia: Wolters Kluwer /Lippincot Williams & Wilkins.
McKee, T., McKee, J., Araiza Martínez, M. and Hurtado Chong, A., 2014. Bioquímica. Las Bases Moleculares De La Vida. 5th ed. México ; Madrid: McGraw-Hill Interamericana.
Fuentes
Gartner, L., 2017. Texto De Histología Atlas A Color. 4th ed. Barcelona: Elsevier
Harvey, R., Ferrier, D. Palacios Martínez, R., 2011. Bioquímica. 5th ed. Philadelphia: Wolters Kluwer /Lippincot Williams & Wilkins.
McKee, T., McKee, J., Araiza Martínez, M. and Hurtado Chong, A., 2014. Bioquímica. Las Bases Moleculares De La Vida. 5th ed. México ; Madrid: McGraw-Hill Interamericana.
No hay comentarios.:
Publicar un comentario